- Живое состояние
- Элементарная живая машина
- Отличие мертвого состояния от активного живого
- Что нам дает пища
- Что выполняет роль проводов в нашем теле
Живое состояние
В теории Линга жизнь и смерть клетки определяются на основе концепции «живого состояния». Поэтому начнем с того, что выясним: живое состояние — что это вообще такое?
Сам термин «живое состояние» использовали время от времени и раньше, но лишь как замену слову «жизнь». Однако были и исключения. Например, идея Аристотеля о жизни как «форме» или организации. В Эпоху Возрождения (с XIV по XVII век) ученые заметили, что в неблагоприятных условиях организм может перейти в состояние спячки (анабиоза), не уходя при этом из жизни. Это также привело их к мысли, что неотъемлемой составляющей жизни является ее организация. Но еще более содержательной была апелляция Жана Батиста Ламарка (1744-1829) к «état de chose» (положению вещей), и понятие Лепешкина — «состояние жизни», появившееся два столетия спустя после Ламарка.
«État de chose» Ламарка, как и «состояние жизни» Лепешкина — предтечи «живого состояния», как оно понимается в теории Линга. Однако живое состояние в теории Линга радикально отличается от более ранних идей тем, что его можно выразить четкими физико-химическими терминами.
Прежде чем перейти к определению живого состояния, как оно видится в теории Линга, нам необходимо выяснить — что есть мельчайшая единица живой материи? Большинство людей, не знающих о кризисе теории мембранных насосов, полагает, что живой может быть только целая клетка, и не менее чем целая клетка. Однако это не так.
Несложно найти признаки жизни и на субклеточном уровне. Так, участок нервного отростка — аксона, отделенный от тела нервной клетки с ядром, никак не может рассматриваться в качестве целой клетки. Но при этом он сохраняет все свои электрические функции, и именно на нем получена основная часть нынешних сведений о функционировании нервного волокна. Как же можно считать его мертвым? Капля эндоплазмы водоросли Nitella может три дня прожить в подходящей среде. А ведь она тоже никак не может считаться целой клеткой.
С точки зрения теории Линга, наименьшая единица живой материи — не клетка, а ее структурный материал — протоплазма. Именно после появления такой концепции живого состояния впервые обрела жизнеспособность сама концепция протоплазмы, выдвинутая еще полтора века назад Дюжарденом, фон Молем и остальными. Конечно, лишь протоплазма, поддерживаемая в живом состоянии, может служить «физической основой жизни», как ярко выразился Томас Хаксли. Смерть протоплазмы — это и есть конец жизни.
Однако необходимо подчеркнуть, что, в контексте теории Линга, структурная организация протоплазмы может принимать самые разные формы. Это не всегда описанное Дюжарденом, фон Молем и другими гистологами вязкое, но текучее «живое желе», которое коллоидные химики называют «гелем» или «коацерватом». Если мы рассечем мышцу лягушки, либо аксон кальмара, вязкая желеобразная консистенция протоплазмы не даст ей вытечь наружу — в противоположность эндоплазме поврежденной клетки водоросли Nitella. Несмотря на различия в консистенции, любая протоплазма обладает способностью существовать в живом состоянии и переходить в мертвое состояние, если к этому принуждают изменения окружающей среды.
Представить физико-химическое определение живого состояния поможет простая модель — цепочка стальных гвоздей, свободно разбросанных на стекле, соединенных друг с другом гибкой проволочкой и посыпанных железными опилками. Очевидно, что цепочка из гвоздей имеет случайную конфигурацию и не взаимодействует с железными опилками. Если теперь снизу поднести к стеклу сильный магнит — все, что лежит на нем, намагничивается и фиксируется. В результате этого простого действия не только цепочка гвоздей приобретает более жесткую и упорядоченную (спрямленную) «конформацию» — намагничиваются также хаотично разбросанные вокруг железные опилки, которые притягиваются к цепочке гвоздей поодиночке или скоплениями, т.е. множеством поляризованных слоев. В итоге образуется единая система из сцепленных гвоздей, железных опилок и магнита, свободная энергия которой выше, чем у исходной, а энтропия (хаотичность, дезинтеграция, разупорядоченность) такой системы ниже, чем у системы невзаимодействующих элементов. Удаление магнита размагничивает гвозди и железные опилки — система распадается, возвращаясь в исходное высокоэнтропийное состояние с более низким уровнем свободной энергии и подверженностью любым изменениям. Первое, низкоэнтропийное называется «живым» (с высокой потенциальной энергией), а второе, высокоэнтропийное — «мертвым» (с низкой потенциальной энергией).
Так вот протоплазма — будь то вязкая жидкость или плотный гель — является тесно ассоциированной и кооперативно связанной системой элементов — белков, ионов и воды. Белок — аналог цепочки гвоздей. Ионы К+ и молекулы воды — железные опилки. И, наконец, аналогом магнита является главный кардинальный адсорбат — АТФ. Протоплазма, поддерживаемая в живом состоянии — подобно модели с магнитом, гвоздями и опилками — имеет низкую энтропию и высокую потенциальную энергию, но в протоплазме вместо намагничивания и размагничивания имеет место электронная поляризация и депо-ляризация (или, проще, индукция).
Живое состояние может быть различным. Протоплазма может существовать в покоящемся живом состоянии или в активном живом состоянии, которые перекликаются соответственно с пассивным и активным состоянием жизни в трудах Лепешкина. В активном живом состоянии потенциальная энергия ниже, а энтропия выше, чем в покоящемся. Переход из покоящегося состояния в активное — суть большинства физиологических явлений, и этот переход, как правило, приводится в действие энтропией. Иными словами, именно прирост энтропии в первую очередь является движущей силой перехода протоплазмы из покоящегося состояния в активное.
Однако утверждение, что движущей силой физиологических процессов является рост энтропии, справедливо лишь для процессов, совершающихся однократно — при переходе из покоящегося состояния в активное. Если же речь идет о циклическом процессе, в результате которого система постоянно возвращается в исходное состояние покоя, энтропия и энергия этой системы остаются неизменными. В этом процессе совершается лишь работа за счет энергии, полученной из питательных веществ.
Предельной альтернативой живому состоянию является мертвое состояние, в котором протоплазматический ансамбль достигает самого низкого уровня потенциальной энергии и крайних, высоких, значений энтропии. Таким образом, движущей силой смерти также является рост энтропии. Вот почему жизнь чрезвычайно чувствительна к превышению определенных температурных пределов. Например, мышца лягушки погибает при 41 °С, хотя эта температура тела обычна для многих птиц.
Все возможные состояния живой системы разделены энергетическими барьерами различной высоты. Эти барьеры исключают спонтанный переход клетки или протоплазмы из одного состояния в другое. Сами эти переходы, как правило, резки и подчиняются правилу «все или ничего». Причину этого мы исследуем в других публикациях.
Элементарная живая машина
В 1939 году Энгельгардт и Любимова взбудоражили мировое сообщество физиологов простым экспериментом. Оказывается, АТФ, конечный продукт метаболизма питательных веществ, вызывает укорочение нагруженной белковой нити, полученной из сократительного белка мышечных волокон — актомиозина. При этом АТФ подвергается гидролизу вследствие АТФазной активности данного сократительного белка. Этот опыт наглядно показал, как скрытая химическая энергия, содержащаяся в высокоэнергетических фосфатных связях АТФ, может переходить в механическую работу.
Однако концепция высокоэнергетической фосфатной связи была опровергнута еще в середине 1950-х — ни одна фосфатная связь АТФ не содержит энергии, которую можно было бы использовать. Вскоре после этого революционного открытия такие ученые, как Райзман и Кирквуд, а также Моралес и Боттс, принялись за поиски иного способа, каким бы АТФ могла осуществлять свою ключевую роль в клетке. В своей книге Линг рассказывает о том, как он однажды использовал идею Райзмана и Кирквуда, чтобы объяснить роль АТФ в избирательном накоплении К+ клетками, и как впоследствии заменил первую модель своей теории другой — индуктивной.
В новой модели к прямому электростатическому эффекту (D-эффекту), распространяющемуся в пространстве согласно закону Кулона (как и в модели Райзмана-Кирквуда), добавлен более прецизионный индуктивный эффект (I-эффект), распространяющийся по цепи атомов. Так как в большинстве физиологических явлений, происходящих в протоплазме, явно преобладает именно индуктивный компонент, имеет смысл продолжать называть индукцией то, что, строго говоря, следовало бы называть прямым F-эффектом, как это делал Линг в своих первых работах (поскольку на практике I- и D-эффекты трудно различать, их суммарное действие обозначается в теории Линга как F-эффект).
На приводимом рисунке изображена схема минимального фрагмента протоплазмы, который обладает всеми необходимыми чертами живого состояния покоя и способен к структурным изменениям, необходимым для перехода в альтернативное, обратимое, активное, живое состояние, и необратимое — мертвое состояние. Иными словами, это схема элементарной живой машины.
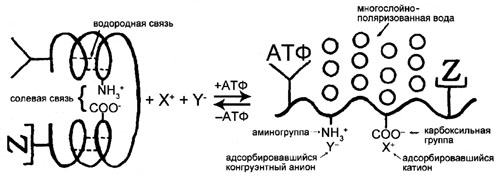
Слева и справа — альтернативные состояния молекулы белка, определяющие альтернативные состояния протоплазмы: состояние покоя (справа) и активное или мертвое состояние (слева). Различия между этими двумя последними состояниями будут рассмотрены ниже. Адсорбция активным белком — его кардинальным АТФ-связывающим центром — кардинального адсорбата АТФ, а также «помощников», в том числе конгруэнтных анионов (показанных здесь как «адсорбировавшийся конгруэнтный анион») и белка X (отмеченного знаком «Z»), разворачивает интровертную (свернутую) вторичную структуру белка. В ходе этой трансформации рвутся солевые связи, и освобождающиеся β- и γ-карбоксильные группы (помеченные в правой части рисунка как «карбоксильная группа») начинают избирательно сорбировать ионы К+, а освободившиеся от участия в формировании вторичных структур NH- и CO-группы пептидных связей начинают взаимодействовать с водой, формируя многослойную структуру из поляризованных и ориентированных молекул воды — одного из ключевых факторов в поддержании живого состояния покоя.
Теоретически, чтобы вдохнуть жизнь в умирающую элементарную живую машину и вернуть ее в покоящееся живое состояние, нужно развернуть интровертную активную либо мертвую протоплазму, и снова превратить ее в экстравертную (развернутую). Для этого нам нужна, кроме АТФ [1], поляризованная и ориентированная вода [2], способная ослабить внутри- и межмолекулярные водородные связи между NH- и CO-группами полипептидного остова и «развернуть» их для взаимодействия с водой. Кроме того, нам нужен К+ [3] для разрыва солевых связей между β- и γ-карбоксильными группами и фиксированными катионами (ε-аминогруппами и гуанидиновыми группами). Но К+ не сможет сделать это один — чтобы помочь К+ ослабить солевые связи, нужны особые «конгруэнтные» анионы [4], способные оттеснять фиксированные катионы и фиксированные анионы друг от друга.
Конгруэнтные анионы в разных клетках — разные. В эритроцитах это, по большей части, Cl-, в мозге и сетчатке морской свинки — L-глутамат, в аксоне кальмара — изетионат, а в мышцах лягушки — органические фосфаты.
Еще одним ценным помощником АТФ является схематично изображенное на рисунке выше вещество (отмеченное знаком «Z»), которое Линг называет белком (или белками) Х [5]. Исследовательская группа Линга в Пенсильванской больнице гналась по пятам за этим неопознанным веществом, содержащимся в эритроцитах. И вот, они уже почти его идентифицировали, как вдруг их лаборатория была закрыта Национальным институтом здоровья и Управлением морских исследований США. Собрание их послушных консультантов (или что там у них было), потребовало прекращения работы Линга. И, несмотря на невероятно любезную и своевременную помощь доктора Рэймонда Дамадьяна и его компании Fonar, группа Линга оказалась лишена средств, необходимых для завершения поисков, — радиоизотопных технологий, высокоскоростных центрифуг, приборов для ионного анализа и т.д. Загадка белка X так и осталась нераскрытой.
Однако есть веские основания полагать, что белком X (или одним из белков семейства X) является актин в мономерной форме или как часть системы микрофиламентов.
Отличие мертвого состояния от активного живого
Согласно теории Линга, движущей силой физиологических процессов и самой гибели клетки является увеличение энтропии, то есть именно рост энтропии является необходимой предпосылкой для перехода системы из покоящегося живого состояния в активное (например, сокращение мышцы). Поэтому нет ничего удивительного в том, что изменения в интенсивно работающей клетке зачастую качественно сходны с изменениями при гибели клетки. Например, повторяющиеся сокращения поперечнополосатой мышцы сопровождаются утратой клеточного К+ и притоком в клетку Na+, что наблюдается и в умирающей мышце. Согласно теории Линга, ключевую роль в развитии обоих состояний играет утрата АТФ.
Многие годы исследователей физиологии мышц озадачивало отсутствие снижения уровня АТФ после одиночного мышечного сокращения. Причиной этому является фермент креатинфосфокиназа, сразу восполняющий затраты АТФ при работе клетки за счет запасов креатинфосфата (КрФ). Когда же Инфант и Дэвис обнаружили, что ядовитое вещество динитрофторбензол (ДНФБ) способно избирательно подавлять креатинфосфокиназу, было сразу же показано, что сокращение мышц, обработанных этим веществом, действительно сопровождается снижением уровня АТФ.
В нормальной мышце запасы АТФ и КрФ постоянно восполняются за счет двух основных метаболических путей: гликолиза и окислительного фосфорилирования. Иодуксусная кислота блокирует этот процесс — поэтому обработанные ею мышцы способны лишь на ограниченное число сокращений. Кроме того, с каждым новым сокращением расслабление отравленной мышцы становится все менее полным до тех пор, пока она полностью не перестает сокращаться, достигнув стойкого укорочения — окоченения, что означает ее гибель.
Итак, главным отличием мертвого состояния от активного живого является необратимость одного и полная обратимость второго, хотя существуют и количественные отличия — степень изменений при мертвом состоянии, как правило, более глубока.
| Главная роль метаболизма — после каждого перехода в высокоэнтропийное состояние с низкой потенциальной энергией обеспечить быстрое и полное возвращение в исходное низкоэнтропийное живое состояние с высокой потенциальной энергией. |
Что нам дает пища
Крайне интересно и поучительно будет узнать, что говорил о жизни один из величайших физиков-теоретиков Эрвин Шрёдингер, один из основателей квантовой механики. В своей книжке «Что такое жизнь с точки зрения физики», вышедшей в 1944 году, Шрёдингер презрел общепринятое убеждение, что мы и все остальные организмы питаемся энергией. Он настаивал: «То, чем кормится организм, — отрицательная энтропия».
Это высказывание вызвало протесты многих ученых, отвечая на которые он признал, что энергетическая составляющая пищи все же имеет значение. Однако теперь мы остались без четкого ответа: чем же мы питаемся — энергией или «отрицательной энтропией»? Почему более чем за полвека, на протяжении которых книжка Шрёдингера переиздавалась не менее 18 раз, никто так и не предложил своего варианта ответа? Мы видим основную причину этой нерешительности в абсурдной теории мембранных насосов, прямо противоречащей предположению Шрёдингера.
Как читатель, вероятно, знает, согласно теории мембранных насосов, клетка содержит лишь свободные вещества и свободную воду, а это — уже максимум энтропии и дальнейший ее рост невозможен. В этом случае живая клетка, согласно теории мембранных насосов, содержит не больше «отрицательной энтропии», чем погибшая. А если учесть, что погибшая клетка склонна к большей жесткости (трупное окоченение), можно даже подумать, что в погибшей клетке больше отрицательной энтропии (упорядоченности), чем в живой. Однако Шрёдингер твердо верил, что именно отрицательная энтропия «бережет нас от смерти».
Неужели мы в самом деле потребляем именно отрицательную энтропию?
И да, и нет. Дело вот в чем. Существуют неопровержимые данные, доказывающие, что адсорбированное состояние клеточного К+ и клеточной воды — неотъемлемая черта покоящегося живого состояния. Адсорбция ограничивает свободу их перемещения и создает тем самым отрицательную энтропию, что полностью согласуется с акцентом Шрёдингера на необходимости высокой отрицательной энтропии для существования жизни. Но нельзя ведь потреблять саму по себе отрицательную энтропию и прямо направлять ее к молекулам воды и ионам К+, к примеру, чтобы они адсорбировались. Однако мы можем добиться этого косвенно.
Благодаря потрясающим достижениям биохимии нам стало известно, что распад каждой молекулы глюкозы до CO2 и воды в результате гликолиза и окислительного фосфорилирования сопровождается синтезом 36 молекул АТФ. Адсорбция АТФ кардинальными центрами основных внутриклеточных белков (например, миозина в мышечных волокнах) влечет за собой тотальную адсорбцию воды и К+ с утратой ими свободы перемещения и соответствующим ростом отрицательной энтропии. (более подробно о том, как АТФ выполняет эту жизненно важную функцию, будет рассказано в других публикациях).
Итак, мы потребляем не отрицательную энтропию как таковую.
| Мы потребляем пищу, содержащую доступную для использования энергию, благодаря чему образуется АТФ. АТФ, в свою очередь, адсорбируясь на белке, вызывает адсорбцию большей части внутриклеточного К+ и, что самое важное, — адсорбцию почти всей внутриклеточной воды с образованием множества поляризованных, ориентированных слоев. |
При этом отрицательная энтропия существенно растет (а по абсолютной величине — снижается) и поддерживается на высоком уровне клетками в состоянии покоя. При циклической работе клетки, когда с каждым новым циклом протоплазма возвращается в исходное состояние покоя, энергия пищи тратится на ресинтез АТФ, затрачиваемой во время каждого цикла физиологической активности для восстановления этого исходного состояния. Мы использовали модель с магнитом, чтобы продемонстрировать механизм дистанционной каскадной регуляции энергии системы и ее энтропии. Однако уже говорилось, что аналогом магниту в живой клетке, как гласит теория Линга, являются внутримолекулярные электронные взаимодействия, которые и будут предметом рассмотрения в дальнейшем.
Электрический импульс, переданный по медному проводу длиной в 7 земных экваторов, пройдет это расстояние менее чем за секунду. Исключительная скорость электрической передачи по проводам, открытая Бенджамином Франклином, позволила создать такие великие изобретения, как телеграф, телефон и интернет, превратившие обычных людей в волшебников, ежедневно творящих технические чудеса.
Естественно, биологи задавались вопросом о том, может ли в основе скоординированности физиологических явлений лежать тот или иной способ дистанционной передачи сигналов. Но ответ оказался отрицательным — нет, не может. Единственным кандидатом на роль биологических проводов могли бы быть вездесущие белки с их длинными молекулами. Но белки не являются проводниками. Наоборот, нити такого типичного белка, как шелк, используют для подвешивания и изоляции металлических шаров при школьных демонстрациях феномена электростатического взаимодействия зарядов.
Конечно, все слышали, что сигналы могут передаваться в виде потенциалов действия по мембране нервных и мышечных волокон. Но мало кому известно, что в основе функционирования элементарной живой машины на молекулярном уровне лежит дистанционный каскадный контроль. Более того, оба эти способа дистанционной передачи сигнала имеют близкий механизм, что можно продемонстрировать на искусственной модели.
Более двух тысячелетий тому назад враждебные взаимоотношения с гуннами (называемыми в Китае Hsiung-nu) привели к возведению Великой китайской стены, которую можно рассматривать как своего рода аппарат для скоростной передачи информации на большие расстояния. Длиной более 6350 км, пересекающая пустыни и скалистые горы, эта рукотворная структура видна даже из космоса. Отлично вымощенная и настолько широкая, что по ней рядом могут скакать две лошади, она являлась также скоростным шоссе древности, поднятым над землей на много метров.
Стена соединяет между собой череду сторожевых башен, расположенных друг от друга на расстоянии 200-500 метров. При появлении неприятеля часовые, находящиеся в такой башне, тут же разжигали костер и подавали этим световой и дымовой сигнал другим постам. Увидев их, часовые соседних сторожевых башен в свою очередь тоже зажигали костры, передавая сигнал тревоги дальше. И так сигнал быстро передавался по цепочке сторожевых башен, пока не достигал гарнизонов, охранявших границу. На помощь постам, подвергшимся нападению, быстро прибывала конница и отражала натиск врага. Этот цепной способ передачи сигналов по Великой китайской стене может служить моделью быстрой передачи информации способом, альтернативным проводной связи — передача информации на большие расстояния обеспечивается многократным повторением ближних взаимодействий соседних сторожевых башен. Эта модель обладает существенным сходством как с распространением потенциала действия вдоль нервных и мышечных волокон, так и с распространением индуктивного эффекта вдоль белковых цепей (согласно теории Линга).
Белки — основной субстрат жизни. Нельзя отрицать важность и ДНК с РНК, без которых жизнь не смогла бы долго продолжаться. Но функции ДНК и РНК не выходят за рамки функции кодирования ими белков (если не считать самокопирования), так что их роль не так тесно связана с жизнью, как роль белков.
Каждый белок — это уникальная последовательность аминокислотных остатков, разнообразие которых, как правило, составляет двадцать две аминокислоты. Поэтому язык жизни можно сравнить с письменным языком (английским, например), алфавит которого включает двадцать шесть букв. Слова еаt и аtе имеют разные значения, хотя набор букв в них одинаков. Так же и белок характеризуется не только природой и количеством аминокислотных остатков, но и порядком их расположения в цепи.
Разница между словами еаt и аtе чисто внешняя. Различие же между разными последовательностями аминокислотных остатков гораздо более глубокое. Во-первых, у них разная форма молекулы, или конформация. Еще важнее их электронные отличия в силу действия индуктивного эффекта.
В качестве примера приведем превращение приятного острого уксуса или уксусной кислоты (CH3COOH) с константой диссоциации 4.75, в чрезвычайно едкую трихлоруксусную кислоту (CCl3COOH) с константой диссоциации < 1.
Каждый атом хлора в молекуле трихлоруксусной кислоты более электроотрицателен, чем замещенный им атом водорода. Понятно, что атом хлора с 17-ю положительно заряженными протонами в ядре сильнее притягивает отрицательно заряженные электроны, чем замещенный им атом водорода с его единственным протоном.
Поэтому при замещении атомов водорода на хлор притяжение к ним электронов возрастает. Более того, эффект притяжения электронов в конце концов достигает по цепочке атомов отрицательно заряженного атома кислорода карбоксильной группы. При этом электронная плотность на этом атоме снижается и связь с положительно заряженным атомом водорода, — протоном (Н+), — ослабевает. При замещении же всех трех атомов водорода на хлор образуется кислота с очень низким сродством к Н+. И слабая кислота в результате индуктивного эффекта становится очень сильной.
Белки — основное вещество протоплазмы, и в природе их умеют создавать лишь живые клетки, включая и те, из которых состоят сами биохимики, научившиеся синтезировать белки искусственно. Уникальными соединениями их делает вовсе не разнообразие аминокислотных остатков. Сколько бы их разновидностей ни было еще открыто, это не изменит главного: по-настоящему уникальной структурой белков является их полипептидная цепь.
Традиционная химия белков не уделяет этому уникальному структурному компоненту любой формы жизни того внимания, которого он заслуживает. А вот теория Линга с самого своего основания придавала полипептидной цепи огромное значение как уникальной резонансной структуре. Имеется в виду кето-енольная таутомерия: динамическое равновесие между кето-формой пептидной связи и енольной (—С=O—NН— ↔ —СОН=N—). Благодаря таутомерии пептидная связь имеет частично (на 40%) характер двойной (сопряжённой) связи, что проявляется в уменьшении её длины (1.32 Å), по сравнению с длиной одинарной —С—N— связи (1.47 Å). Эта «вибрация» делает длинную полипептидную цепь высокополяризуемой и, тем самым, придает ей способность передавать индуктивный эффект на большие расстояния. Эта особенность сближает цепь пептидных связей с Великой китайской стеной, а также с более непритязательной цепочкой из гвоздей, превращая пептидный остов белка в главную информационную трассу элементарной структуры жизни.
Как же далеко может передаваться локальный или, как его называет Линг, прямой индуктивный эффект по полипептидной цепи белка in vitro? Существует, по меньшей мере, четыре группы независимых экспериментальных данных, дающих ответ на этот вопрос. Взятые вместе, они показывают, что прямой индуктивный эффект может передаваться через три пептидные связи, и еще — через короткий участок насыщенной метиленовой цепи бокового радикала. Если вернуться к аналогии с Великой китайской стеной, то дистанцию, на которую распространяется индуктивное влияние, можно уподобить расстоянию между двумя ближайшими сторожевыми башнями.
Ввиду относительно быстрого ослабления индуктивного эффекта метиленовыми группами, функциональные группы, расположенные на коротких боковых радикалах, в большей мере испытывают его влияние, чем более удаленные. В числе этих проксимальных функциональных групп находятся, прежде всего, пептидные связи полипептидного остова белка и уже знакомые нам β- и γ-карбоксильные группы. Кроме того, ближе других к остову белка расположены функциональные группы на коротких боковых цепях остатков серина, треонина, цистеина, тирозина, фенилаланина, гистидина и триптофана. Так как функциональная группа каждого из них отделена от полипептидного остова не более чем двумя метиленовыми группами, индуктивному эффекту не так сложно достичь их. |
